2020年 | プレスリリース・研究成果
シロイヌナズナが自殖へと進化した仕組みを解明 花粉遺伝子に生じた二重変異のシナジー効果による二段階での制御メカニズム
【発表のポイント】
- 遺伝的多様性を維持する仕組みとして、植物は他殖性を発展させてきた。
- シロイヌナズナの他殖性から自殖性への進化は、自家不和合性を制御する花粉遺伝子SCRに生じた逆位と転写量低下を引き起こす変異の両方が原因。
- 高等植物の自殖と他殖を自由に制御できる可能性を開いた。
- アブラナ科野菜の効率的な品種改良への応用が期待される。
【概要】
植物の生殖過程には自己・非自己の花粉を認識することで子孫を残すべき花粉を選別する「自家不和合性*1」という仕組みがあります。シロイヌナズナは進化の過程でこの自家不和合性を失い、自身の花粉で子孫を残す「自家和合性」の種(しゅ)へと変化してきましたが、その詳細な進化の道筋は不明でした。
東北大学大学院生命科学研究科の渡辺正夫教授らの研究グループは、三重大学、テキサス工科大学、明治大学、東京大学、チューリッヒ大学、横浜市立大学、大阪教育大学との共同研究により、植物研究のモデル生物であるシロイヌナズナが自身の花粉で子孫(種子)を残す「自殖」へと進化した原因が花粉で機能するたった1個のSCR*2遺伝子の2種類の変異にあり、ゲノム情報と転写の二段階による変異のシナジー効果*3で制御されていることを明らかにしました。
本成果は、2020年9月11日、スイス科学誌「Frontiers in Plant Science」(電子版)に掲載されました。本研究は文部科学省科学研究費補助金、日本学術振興会科学研究費、スイス科学基金の支援を受けて行われました。
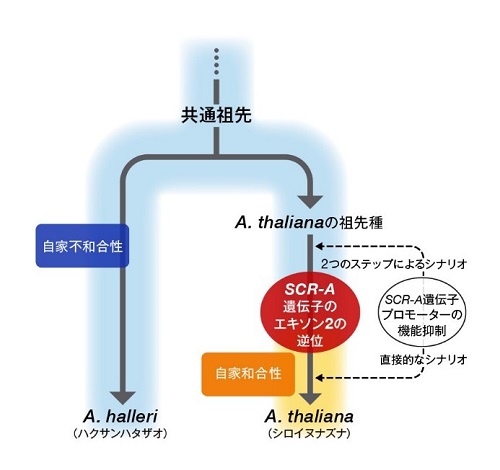
図. シロイヌナズナの自殖への進化モデル図
アブラナ科共通祖先種は自家不和合性を有しており、そこから分岐したハクサンハタザオ(A. halleri)は現在においてもそれを受け継ぎ維持している(図左側)。それに対して、シロイヌナズナ(A. thaliana)は、進化の過程においてSCRタンパク質をコードするエキソン領域と転写の制御領域にそれぞれ変異が生じ、自家不和合性を失い自殖性になったと考えられる(図右側)。
【用語解説】
*1 自家不和合性:近親の交雑を続けることによる個体の弱体化を防ぎ、集団の均一化を避けるための機構。雌雄が正常であるにもかかわらず、自己の花粉を認識・拒絶することで、受精には至らない現象。アブラナ科植物では、自己認識を司る因子が♀側・♂側ともに明らかになっており、それぞれ、受容体型キナーゼ・リガンドタンパク質からなる。これらは、個体ごとに構造の異なる多型性を有しており、♀側因子(受容体型キナーゼ)と♂側因子(リガンドタンパク質)が同一個体由来であった場合には、互いに結合することで、めしべ細胞内に自己花粉拒絶のシグナルを伝えることができる。逆に♀側因子と♂側因子が異なる場合には、これらが結合できないために、受粉・受精が正常に行われ、種子を作ることができる。
*2 SCR:アブラナ科自家不和合性の♂側原因タンパク質。花粉表面に付着した小型のタンパク質で、受容体である♀側原因タンパク質SRKと結合する。SP11とも呼ばれる。
*3 シナジー効果:自家和合性に変異する過程において、SCR遺伝子の「エキソン2領域の逆位」と「転写を司る制御領域」が、相乗的に機能することで、自家和合性形質を安定させている。このSCR遺伝子で起きた2つの変異で生じる効果をシナジー効果と呼んでいる。
問い合わせ先
(研究に関すること)
東北大学大学院生命科学研究科
担当 渡辺 正夫 (わたなべ まさお)
電話番号: 022-217-5681
Eメール: nabe*ige.tohoku.ac.jp(*を@に置き換えてください)
(報道に関すること)
東北大学大学院生命科学研究科広報室
担当 高橋 さやか (たかはし さやか)
電話番号: 022-217-6193
Eメール: lifsci-pr*grp.tohoku.ac.jp(*を@に置き換えてください)